We are looking for excellent researchers, at any level of their carrer (undergraduate, master student, PhD applicants, Postdoctoral fellow). We have presently several openings at the PhD and postdoctoral levels, and in case you are interested by our approach do not hesitate to contact us to know more about our projects. If we are interested by your profile, we will support your application to various individual fellowships that are available in France and Europe (e.g. HFSP, Marie Curie Fellowships) at both the Doctoral and Postdoctoral level.
Student Internships
We are also looking for master students that want to join our group for an internship (3-6 month). We'll be happy to train physcisists to biology and biologists to physics and computational biology. You can join us at any time of the year and help us advance anyone of the subject described in this webpage. During your internship, you will have the opportunity to learn several methods and techniques among, fluorescence microscopy, automated microfluidics, image analysis, single cell tracking, cell culture, synthetic biology, optogenetics, mouse embryonic cell culture, molecular cloning, embryology, mathematical modeling of signaling pathway, mechanics of locomotion, collective behaviours ...
Systems Biology of the adaptation to stress of the Yeast S. Cerevisiae.
Cells have evolved complex signaling pathways to cope with environmental stress. Quantitative biology and interdisciplinary approaches can be used to probe the dynamics of signaling pathways in living cells. We studied the signal processing ability of yeast cells in response to osmotic stress ...[+]
Optogenetic control of gene expression and Cell-computer Interfaces
Time lapse fluorescent imaging of cells exposed to time varying stimulus can be used to quantitatively probe the dynamical behavior of such regulatory motifs. In such an experimental framework, cells are often seen as complex machines, which response functions can be measured by time varying stimulations...[+]
How cells encode differentiation signals?
During embryonic development, cells gradually go from pluripotent to highly specialized states. This process is guided by morphogens, a class of secreted factors that have been shown in vitro to be able dictate cell fate in a dose-dependent manner. How morphogens operate in vivo to define cell fate is still unclear...[+]
Morphogenesis and Fitness of yeast colonies.
Microorganisms growing on solid media can form various complex shapes, yet to be fully understood. In our lab, we study Saccharomyces cerevisiae colonies growth by restraining access to nutrients ...[+]
Bottom-up Approach to Mammalian Gastrulation
One of the most striking feature of embryonic development is that differentiation is happening in a spatially ordered fashion: tissue self-organize to form well-defined patterns. During gastrulation, the cells of the embryo are...[+]
Systems Biology of the adaptation to stress of the Yeast S. Cerevisiae.
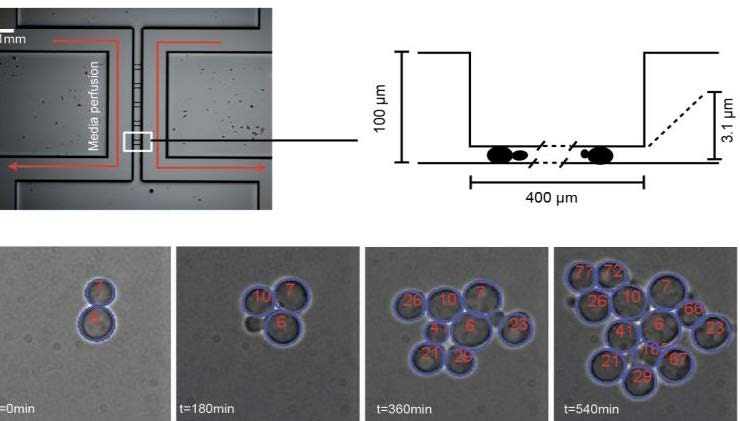
This two-layer microfluidic device can be used to grow yeast cells as a monolayer for very long time. It allows to segment and track yeast cells and to access to single cell gene expression. This allows to perform single cell quantitative analysis and models of gene expression.
We are interested in understanding how simple organisms, such as yeast cells, detect and adapt to changes of their environment. We combine molecular biology, microfluidics and microscopy to observe living cells while they respond to controlled, periodic fluctuations of their chemical environment. We are now able to follow single cells over several divisions while changing their environment at will. We are presently looking for a student who wants to study signaling dynamics and transcriptional adaptation mechanism in fluctuating environment. Among several possible projects, we propose to start by focusing on the cell cycle regulation in presence of an external (osmotic) stress and on the physical limits of gene expression of yeast cells in severe stress conditions.
How is the cell cycle regulated by periodic osmotic stress in yeast cells? Osmotic stresses are known to trigger the cell cycle arrest in G1 or G2 phases, after the activation of the HOG cascade, a very well known MAPK cascade in yeast. The dynamics of this cell cycle arrest have not yet been studied, although it is likely that arrest and recovery of the cell cycle progression should be done with a correct timing. Here we propose to subject yeast cells to periodic osmotic stress using micro fluidics and to monitor cell divisions through several generation over a range of period and stress intensity. We will reconstruct the lineage of cells and observe the effect of genetics modifications of the HOG cascade (mutation, controlled expression through inducible promoters) on the cell cycle arrest duration. This will give us a measurement of the cut off frequency of the different molecular actions that are used by cells to arrest the cell cycle.
How do yeast cells adapt to very high osmotic stress. Yeast cells are known to be able to survive to osmotic stress, by activating the HOG cascade and a large set of genes. However, recent work in our lab show that both this signaling cascade dynamics and transcriptional adaptation of key genes are slowed down for too intense osmotic shock. It can be delayed by several hours. During that time, not much is known on what the cell does. The goal of this project is to study the transcription dynamics of several key genes by fluorescence microscopy, when they are subject to both a strong hyper osmotic shock and a secondary stress (Calcium, Oxidation, Glucose repression, UV…). The differential dynamics of the transcriptional responses to this secondary stress will set the basis to elucidate how cells can adapt and survive to very strong stresses and what are the physical constraints that are placed on these processes
References :
A. Miermont, F. Waharte, S. Hu, S. Bottani, M. McClean, S. Léon and P. Hersen*, Severe osmotic compression triggers a slowdown of intracellular signaling, which can be explained by molecular crowding; PNAS, 110 (14), 5725-30 (2013).
Hersen, P., et al., Signal Processing by the HOG MAP Kinase pathway, accepted in PNAS, 2008.
Optogenetic control of gene expression and Cell-computer Interfaces
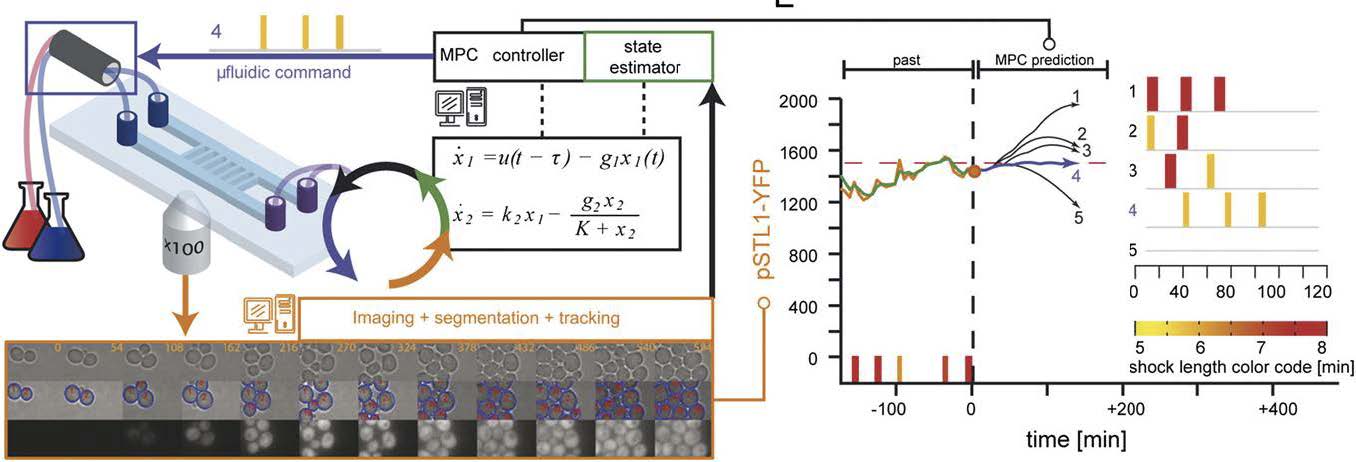
Real time control platform16. It is composed of a microscopy system, a micro-fluidic device to change in real time the extracellular environment (left), and a set of algorithms to segment the images, measure the fluorescence of every cells (bottom) and estimate the deviation from the desired real time profile of gene expression (right). Different control strategies and state estimation have been tested successfully. For the CoGEx Project, we will upgrade this system into a versatile platform for the control of population of cells or single cells using chemical or light inducible gene expression systems.
Time lapse fluorescent imaging of cells exposed to time varying stimulus can be used to quantitatively probe the dynamical behavior of such regulatory motifs. In such an experimental framework, cells are often seen as complex machines, which response functions can be measured by time varying stimulations, as it is classically done in electrical and mechanical engineering. Several recent studies have used such approaches to constrain the modeling of regulatory gene networks and signaling pathways. We are, however, far from being able to construct models of biological processes which are as predictive and robust as it is usually the case in physics and engineering. One of the main difficulties is the limited knowledge of the cell state (metabolism, cell cycle, size, age…) that can be accessed through fluorescence imaging and the existence of noise associated with gene transcription. In turn, this strongly limits our ability to drive cellular processes, such as gene expression, over long time with a quantitative accuracy. Nevertheless, having a mean to externally control, in real time, the expression level of a gene of interest, and use this to generate time varying perturbations from within a regulatory network would be a major step towards a better, quantitative understanding of how a cell functions. This would also have important consequences for applied biotechnology. Indeed, a major challenge of synthetic biology is to engineer cells that can robustly perform a synthetic program in a broad range of environmental conditions and despite the stochastic nature of gene expression.
This motivated us to develop a real time, computer based feedback loop control of gene expression in yeast cells. The principle of controlling a dynamical system thanks to a feedback loop has been used extensively in engineering and is a key feature of most electromechanical tools of our everyday life. The basic idea is simple: monitor the readout and operate a change on the system to adjust it in real time so that it follows a given target profile. This permits to compensate for environmental fluctuations and un-modeled dynamics. Implementing such strategy in biological systems turned out to be a challenge that we and two other teams recently solved (Ulhendorf et al. 2012, Milias-Argeitis et al. 2011, Toettcher et al. 2011). In particular, we were the first to successfully force the level of expression of a fluorescent reporter gene in yeast to follow a time varying profile at the population and at the single cell level over multiple cell generations. This puts us in a unique position to develop experimental systems aiming at controlling cellular processes through cell-computer interfaces.In this context, we are now further developing the experimental and theoretical tools for the computer-based remote-control of live cells. We are looking for a M2 internships to help us improve our open platform based so that we can use optogenetic for the control of gene expression in yeast and/or the control of signaling pathways in mammalian cells. This work will be done in close collaboration with G. Batt (INRIA) with whom we have a long lasting collaboration.
J. Uhlendorf, A. Miermont, T. Delaveau, G. Charvin, F. Fages, S. Bottani, G. Batt*, P. Hersen*, Long-term model predictive control of gene expression at the population and single-cell levels; PNAS, 109 (35), 14271-6 (2012)
How cells encode differentiation signals?
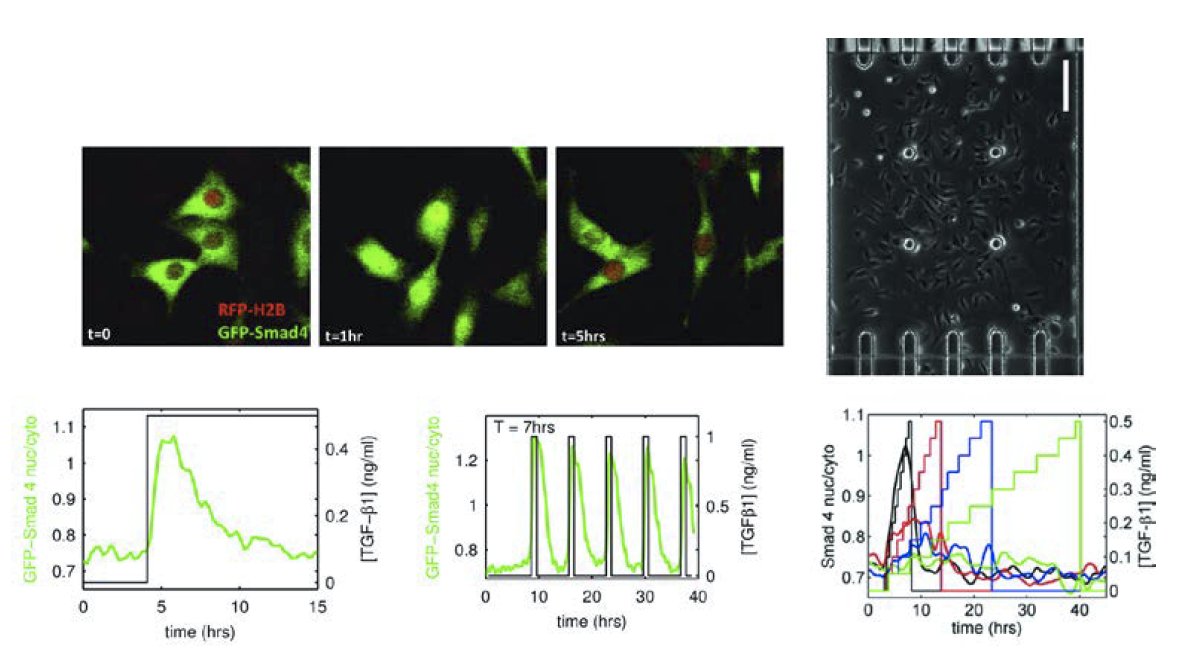
top : example of fluorescent reporter for TGF-β signaling (green): the reporter is in the cell nucleus when the pathway is active. Bottom: example of cell response to various temporal profiles of stimulation. From left to right: step, pulses and ramps at various speeds. The response to a TGF-β concentration step is transient and adaptive, slowly increasing the ligand concentration diminishes the response and well-spaced pulses of ligand combine additively resulting in greater pathway output than with constant stimulation
During embryonic development, cells gradually go from pluripotent to highly specialized states. This process is guided by morphogens, a class of secreted factors that have been shown in vitro to be able dictate cell fate in a dose-dependent manner. How morphogens operate in vivo to define cell fate is still unclear, especially as we have recently shown that the temporal profile of the concentration of morphogen could be as important as its absolute concentration. (see Sorre et al. 2014)
We will focus on Nodal signaling, a morphogen of the TGF-β family that plays a crucial role in early development as well as in cancer. Using a combination of automated microfluidic cell culture and single cell analysis, we propose to study how mouse embryonic stem cells encode Nodal signaling and how this defines their subsequent differentiation into ectoderm, mesoderm or endoderm.
The project will be done in close collaboration with the lab of J. Collignon at IJM and could be extended to a PhD thesis project.
Sorre, B., Warmflash, A., Brivanlou, A. H., & Siggia, E. D. (2014). Encoding of Temporal Signals by the TGF-β Pathway and Implications for Embryonic Patterning. Developmental Cell
Warmflash, A., Sorre, B., Etoc, F., Siggia, E. D., & Brivanlou, A. H. (2014). A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nature Methods
Warmflash, A., Zhang, Q., Sorre, B., Vonica, A., Siggia, E. D., & Brivanlou, A. H. (2012). Dynamics of TGF-β signaling reveal adaptive and pulsatile behaviors reflected in the nuclear localization of transcription factor Smad4. PNAS
Morphogenesis and Fitness of yeast colonies.
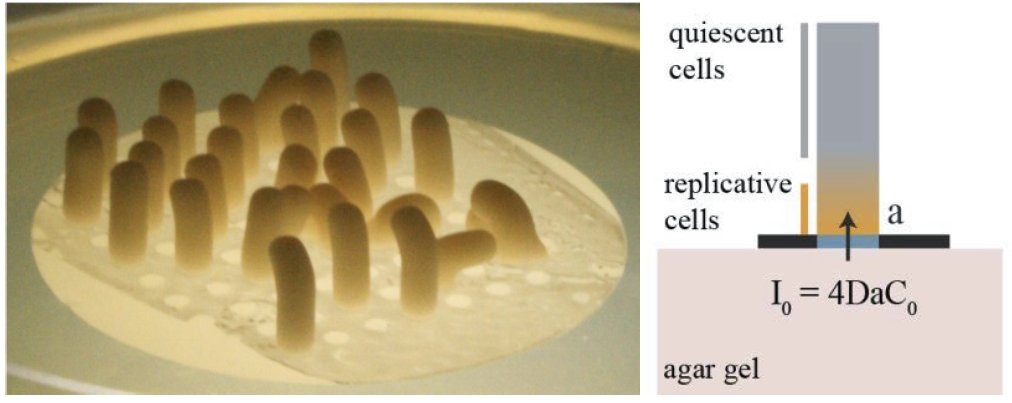
(left) An array of yeast cylindrical colonies. Using a laser based scanner, we can measure the morphogenesis of yeast colonies with an excellent accuracy (< 10 µm). Using this we can relate the growth rate to the incoming nutrient flux (right) and the geometry of the colony.
Microorganisms growing on solid media can form various complex shapes, yet to be fully understood. In our lab, we study Saccharomyces cerevisiae colonies growth by restraining access to nutrients using patterned filtration membrane with selectively blocked pores. When grown on such membranes, yeast colonies can adopt any shapes, growing as if they were extruded from the patterns of porisity. In particular, it is possible to grow yeast cylindrical colonies which grow linearly with time for typically two weeks. This growth rate can be predicted assuming one limiting nutrient diffusion (glucose) and that only some cells near the bottom of the cylinder (closer to the glucose source) are able to divide. Importantly, we showed that the growth rate of the cylinder, does not depend only on the single cell growth rate, but is rather a function of the the single cell growth rate and the specific absorption of glucose. This suggests that cells that are the fittest in well mixed, liquid culture, are not necessarily forming the fastest growing colonies.
To check this hypothesis, we are looking for a M2 physicist to improve our prototype of a colony scanner in order to automatically follow the growth rate of yeast colonies on different carbon sources and with different genetic background. These results will be compared to the growth rate of single cells grown in similar conditions. This way we will compare the "fitness" of an assembly of cells to the fitness of its individuality and outline the emergence of complex, collective behaviour that are set by yeast metabolic interactions with their environment.
C. Vulin, J-M. Di Meglio, A. B. Lindner, A. Daerr, A. Murray, P. Hersen, Growing Yeast into Cylindrical Colonies, Biophysical Journal, 106, 2214–2221, 2014.
Hallatschek O., Hersen P., Ramanathan S., Nelson D., Genetic drift at expanding frontiers promotes gene segregation; PNAS, 104, 19926-19930 (2007).
Bottom-up Approach to Mammalian Gastrulation
ESCs grown under standard and micropatterned conditions show heterogeneous and standardized colony geometries, respectively. Reproducible spatial patterning observed after 42h of differentiation in BMP4 (Sox2: ectoderm, Bra:mesoderm, Cdx2 :trophectoderm).
One of the most striking feature of embryonic development is that differentiation is happening in a spatially ordered fashion: tissue self-organize to form well-defined patterns. During gastrulation, the cells of the embryo are allocated into three germ layers: ectoderm, mesoderm and endoderm. Elegant work in the fish, frog, and mouse embryos has established that spatial patterning during gastrulation is under the control of the morphogen pathways (Activin/Nodal, BMP, Wnt…). However, studying the spatio-temporal dynamics of pattern formation is difficult in those systems, because of their inherent lack of observability (especially in organisms that develop in utero) and manipulability: it is not possible in an embryo to control in a quantitative manner the parameters that are likely to be relevant for the establishment of the multi cellular pattern such as the size and shape of the tissue and its physical and chemical environment. Recently, we took the first step toward in vitro recapitulation of early embryonic patterning by showing that Embryonic Stem Cells (ESCs) confined to circular disks comparable in size to mammalian embryos using micropatterning technology and treated with the gastrulation inducing signal BMP4 differentiate to an outer trophectoderm-like ring followed by the three embryonic germ layers in an ordered, reproducible sequence along the radial axis of the colony, see Fig.1. These results demonstrate that geometric confinement is sufficient to trigger self-organized patterning in hESCs and that the intrinsic tendency of stem cells to make patterns can be harnessed by controlling colony geometries, and provide a quantitative assay for studying paracrine signaling in early development.
The observed pattern doesn’t however recapitulate all the features of the patterning observed in embryos, suggesting that the imposed boundary conditions are too stringent incorrectly defined. By taking advantage the micro-fabrication and the micro-fluidics toolbox, we propose here to study how the physical (stiffness of the substrate, size and shape of the tissue) and chemical (spatio-temporal profile of differentiating signals) properties of the cells micro-environment is affecting the final pattern, in order to define what is the minimal set of boundary conditions allowing for proper gastrulation.
During this internship, you will have the opportunity to learn the following techniques/concepts: micro-fabrication, micro-patterning, surface modification for cell culture, image analysis, single cell tracking, mouse Embryonic Stem Cell culture, molecular cloning, classical embryology, mathematical modeling of cell fate decision and patterns formation.
The project will be done in close collaboration with the lab of J. Collignon (mouse embryology) at IJM and could be extended to a PhD thesis project.
Sorre, B., Warmflash, A., Brivanlou, A. H., & Siggia, E. D. (2014). Encoding of Temporal Signals by the TGF-β Pathway and Implications for Embryonic Patterning. Developmental Cell
Warmflash, A., Sorre, B., Etoc, F., Siggia, E. D., & Brivanlou, A. H. (2014). A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nature Methods
Warmflash, A., Zhang, Q., Sorre, B., Vonica, A., Siggia, E. D., & Brivanlou, A. H. (2012). Dynamics of TGF-β signaling reveal adaptive and pulsatile behaviors reflected in the nuclear localization of transcription factor Smad4. PNAS
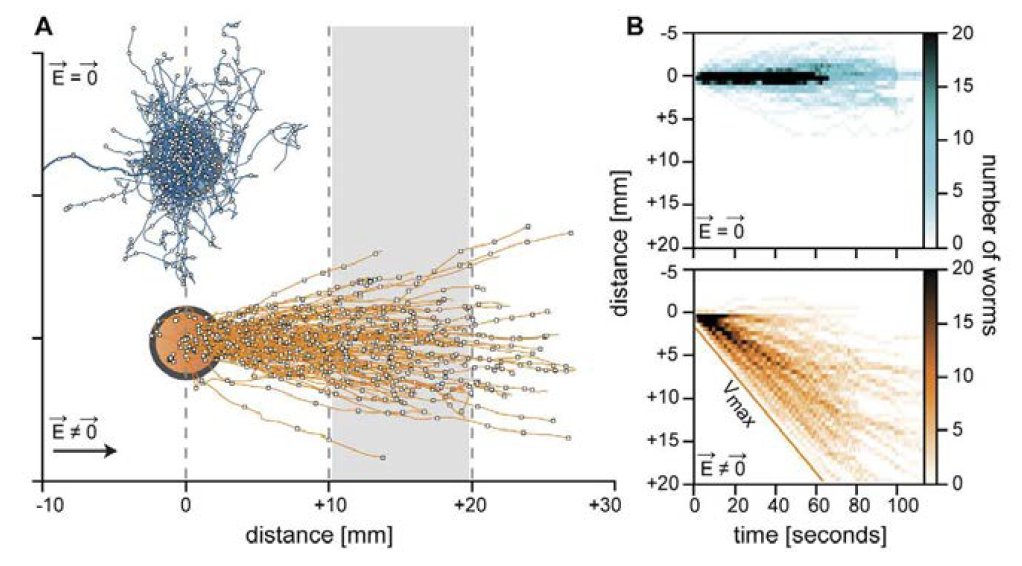
Robustness of electrotaxis locomotion in C. elegans
Typical trajectories of C. elegans worms without (blue) or with (orange) the presence of an electric field. The electrotactic behaviour is very robust for a constant electric field and can be used to serial sort worms according to their locomotor abilities.
Worms (C. elegans) have the striking capacity to detect electric field and navigate towards low potential regions. Here, we want to address how robust this sensory behavior is depending on the electric field properties. We have built an automated experimental platform that allows us to apply an electric field and follow the locomotion of single worms while they try to move towards the negative pole. Interestingly after changing the direction of the electric fields several times, some worms lose track of the correct direction and need some time without any stimulation before being able to detect again the presence of an electric field. We want to study this phenomenon quantitatively to find out what sets the robustness of the electrotactic behavior in C. elegans. We will use several mutant strains and advanced image analysis to quantify the trajectories of worms moving in a time varying electric field.
F. Lebois, P. Sauvage, C. Py, O. Cardoso, B. Ladoux, P. Hersen, J-M. Di Meglio, The C. elegans gait is continuously variable and determined by ambient mechanical stress; Biophysical Journal, 102, 2791-98 (2012).
Xavier Manière, Félix Lebois, Ivan Matic, Benoit Ladoux, Jean-Marc Di Meglio and Pascal Hersen., Running worms: C. elegans self-sorting by electrotaxis; PLoS One, 6(2), e16637 (2011).